
Page 1
How colorectal cancer arises and treatment strategies, based on complexity theory
Nat Pernick, M.D.
Last revised 1 June 2020
Presented at ICCS 2020, the Tenth International Conference on Complex Systems, July 2020 (virtual conference).
Abstract
Introduction: Colorectal cancer is the second leading cause of US cancer death after lung cancer, with 53,200
projected deaths in 2020.
Design: We initially review colorectal cancer risk factors, their population attributable fraction (PAF) and their
mechanism of action. We then categorize them within the context of nine chronic stressors previously identified as
causing most adult cancer: chronic inflammation, carcinogen exposure, reproductive hormones, Western diet, aging,
radiation, immune system dysfunction, germ line changes and random chronic stress or bad luck. We then theorize
how colorectal cancer arises and propose treatment strategies based on a complexity theory perspective.
Results: The PAFs for US colorectal cancer risk factors are: nonuse of screening 22%, physical inactivity 16%,
excess weight 10-20%, tobacco 10%, alcohol 10%, Western (proinflammatory) diet 5% and germ line / family history
2-4%. The PAF is unknown or lacks consensus regarding aging, asbestos, diabetes, inflammatory bowel disease and
the protective effects of menopausal hormones and aspirin. The PAF is estimated at <5% for random chronic stress
or bad luck. These risk factors operate through chronic inflammation (excess weight, physical inactivity, tobacco use
and diet, antagonized by aspirin), carcinogen exposure (alcohol, tobacco, diet, asbestos), aging, immune system
dysfunction and germ line changes. We theorize that in the correct cellular context and in the presence of other
chronic stressors, these risk factors promote network changes that reinforce each other within and between colonic
epithelial cells, leading to intermediate (premalignant) and malignant states which ultimately propagate systemically.
Conclusions: No single treatment modality for colorectal cancer is likely to be curative due to its diverse origins and
because aggressive tumors and widespread disease are accompanied by systemic changes different in character
from those present in tumor cells. To attain high cure rates, we propose combining treatment strategies that: (1) kill
tumor cells via multiple, distinct methods; (2) move tumor cells from "cancer attractor" network states towards more
differentiated or less hazardous states; (3) target different aspects of the microenvironment nurturing the tumor; (4)
counter tumor associated immune system dysfunction; (5) identify, reduce and mitigate patient-related chronic
stressors; (6) eliminate premalignant lesions through more effective screening; (7) identify and target germ line
changes associated with tumor promotion and (8) promote overall patient health.
This research was entirely self funded.
There are no conflicts of interest.
Related papers are available at http://www.NatPernick.com
Introduction
This paper discusses how colorectal cancer arises based on complexity theory. We previously reviewed complexity
theory and how it relates to cancer (Pernick 2017a, Pernick 2018a), proposed that chronic cellular stress is the
underlying cause of most adult cancer (Pernick 2017b), and discussed how lung cancer arises based on complexity
theory (Pernick 2018b-Session 400, poster 36, Abstract, Poster).
We discuss the risk factors of colorectal cancer, their population attributable fraction and their mechanism of action
within the context of the chronic stressors. We then discuss novel treatment strategies based on our theory that
chronic disturbances in an interactive web of networks cause and maintain colorectal carcinogenesis.
Colorectal cancer is the #2 cause of US cancer death after lung cancer, with 53,200 projected deaths in 2020 (8.8%
of total cancer deaths; men 28,630, women 24,570, Cancer Facts & Figures 2020). It is the fourth most commonly
diagnosed non skin cancer in the US (after breast, lung and prostate cancer) with a projected 147,950 new cases in
2020 (colon 104,610, rectum 43,340). From 2007 to 2016 there was an annual decline in incidence of 3.6% in adults
age 55 or older but an annual increase of 2% in those younger than age 55. Similarly, from 2008 to 2017 the death
rate for colorectal cancer declined by 2.6% per year in those age 55 or older but increased by 1% per year in adults
younger than age 55 (ibid; see also Virostko 2019). Overall the colorectal cancer death rate in 2017 (13.5 per
100,000) was less than half of that in 1970 (29.2 per 100,000), which is attributed to increased screening, reduced
incidence and improvements in treatment.
Some authors believe that rectal cancer (defined as arising within 15 cm of the anal sphincter) and colon cancer are
different diseases based of differences in molecular carcinogenesis, pathology, surgical topography and treatment
(Paschke 2018, Li 2009), but we follow the American Cancer Society in combining deaths because many deaths

Page 2
from rectal cancer are misclassified as colon cancer (Cancer Facts & Figures 2020). Tumors on the right and left
sides (proximal and distal to the splenic flexure, respectively) also exhibit distinct molecular characteristics and
histology (Baran 2018). Right sided tumors typically have flat histology, mutations in the DNA mismatch repair
pathway and do not respond well to conventional chemotherapy but have more promising results with
immunotherapy. Left sided tumors typically are polypoid, have mutations in the chromosomal instability pathway
(KRAS, APC, PIK3CA and p53 genes), benefit from 5-fluorouracil based regimes and anti-epidermal growth factor
receptor (EGFR) therapy and have a better prognosis.
Population attributable fraction
The population attributable fraction (PAF) is the projected reduction in death or incidence if a risk factor is not present.
PAFs often overlap and add up to more than 100 percent (World Health Organization - Metrics: Population
Attributable Fraction (PAF), accessed 27May20). PAFs are affected by relative risks assigned and may vary greatly
by geographical region due to differences in prevalence of risk factors. For Table 1, we tried to use results common to
several studies but focusing on the US; when studies had variable PAFs, we used the lower figures.
————————————————————————————————————————————
Table 1 - Population attributable fraction of colorectal cancer incidence (United States)
Nonuse of screening 22%
Physical inactivity: 16%
Excess weight (BMI of 25+): 10-20%
Tobacco: 10%
Alcohol: 10%
Diet: 5-10%
Germ line / family history: 2-4%
Random chronic stress or bad luck <5%
Risk factors with no determined population attributable fraction or with no consensus:
Aging, type II diabetes, inflammatory bowel disease, asbestos exposure
Protective effects: low dose aspirin, menopausal hormone therapy
References are available in the text
————————————————————————————————————————————
What causes colorectal cancer
We previously identified nine chronic cellular stressors that commonly cause adult malignancy: chronic
inflammation (due to infection, infestation, autoimmune disorders, trauma, excess weight, diabetes and other
causes (Pernick 2020a, paper); exposure to carcinogens; reproductive hormones; Western diet (high fat,
low fiber, low consumption of fruit and vegetables); aging; radiation; immune system dysfunction; germ line
changes and random chronic stress or bad luck. (Pernick 2017b). These chronic stressors, triggered by risk
factors discussed below, initially cause local network changes. In the correct cellular context and in the
presence of other chronic stressors, these network changes may reinforce each other and propagate within
and between cells, promoting mutational rewiring of networks that produces intermediate (premalignant)
states and ultimately malignancy. In Table 2, we relate the traditional colorectal cancer risk factors to these
chronic stressors. Local network changes interact with the microenvironment and may ultimately disrupt
networks systemically (Shaath 2019), leading to tumor immune tolerance and growth of metastases.
————————————————————————————————————————————
Table 2 - Chronic stressors and colorectal cancer risk factors
Chronic inflammation: diabetes, diet, inflammatory bowel disease, excess weight, physical inactivity, tobacco;
antagonized by aspirin
Carcinogen exposure: alcohol, asbestos, diet, tobacco
Reproductive hormones: alcohol, excess weight; antagonized by menopausal hormones
Western diet: see diet below; also alcohol, diabetes, excess weight, physical inactivity
Aging: see aging below; also diabetes
Radiation: not related to any risk factors
Immune system dysfunction: alcohol, diet, excess weight, tobacco
Germ line changes: see germ line section; also diet, tobacco
Random chronic stress or bad luck: not related to any risk factors
————————————————————————————————————————————
We discuss below the risk factors for colorectal cancer in declining order of population attributable fraction.

Page 3
Colorectal cancer due to nonuse of screening
The largest risk factor for colorectal cancer is nonuse of screening. US screening participation remains suboptimal,
particularly among underserved populations including the uninsured, recent immigrants and racial/ethnic minority
groups (Gupta 2014).
Colorectal cancer results from complex interactions between inherited, environmental and lifestyle factors to produce
adenomas, which may subsequently develop into carcinoma (Muto 1975, Colorectal Cancer Screening (PDQ®)
Health Professional Version, accessed 28May20). Polyp growth may vary based on a punctuated equilibrium model
(long periods of stasis punctuated by rapid periods of transformation and molecular changes), consistent with our
complexity theory approach (Sievers 2017).
Screening which removes adenomas reduces the incidence and mortality of colorectal cancer (Winower 1993,
Brenner 2014, Niikura 2017), although cancer may arise from fast growing lesions that do not pass through an
adenomatous phase or from incompletely removed adenomas (Samadder 2014). The American Cancer Society has
declared an 80% screening rate as a national goal (Cancer.org, accessed 28May20), which appears feasible based
on compliance rates of 80% in integrated health care delivery systems with aggressive reminder programs, such as
Kaiser Permanente in Northern and Southern California (Lee 2013, Li 2018). Meester estimates that an 80%
compliance rate would reduce colorectal cancer incidence by 22% and deaths by 33% (Meester 2015).
Screening options are high sensitivity-guaiac based fecal occult blood test annually, fecal immunochemical test
annually, multi-target stool DNA test every 3 years, computed tomography colonography every 5 years, flexible
sigmoidoscopy every 5 years and colonoscopy every 10 years (Wolf 2018). Screening is now recommended starting
at age 45 years (Mannucci 2019).
The prominence of nonuse of screening as a risk factor for colorectal cancer demonstrates the folly of seeking “a
world without cancer,” the mission of the American Cancer Society. Cancer is a predictable disease in multicellular
organisms (Trigos 2018). We can prevent some cases, detect it sooner and treat it more effectively, but it will always
be part of human life.
Colorectal cancer due to physical inactivity
The population attributable fraction for colon cancer in the US due to physical inactivity was 16.7% in the only
applicable study (Islami 2018). Although similar to Canada (16.1%), this PAF is higher than that for other Western
countries (Australia: 4.8%, Europe: 6%, Germany 11.3%, New Zealand 4.4%, UK: 4.6%, see Table 3).
————————————————————————————————————————————
Table 3 - Population attributable fraction of colorectal cancer incidence due to physical inactivity by country
US (2014, colon only): total 16.7%, men 16.0%, women 17.3% (Islami 2018, table 5)
Australia (2010): total 4.8%, men 4.2%, women 5.4% (Whiteman 2015)
Brazil (2012, colon only): total 6.0%, men 5.7%, women 6.2% (Rezende 2018)
Canada (Alberta, 2012): total 16.1% (Grundy 2017b)
China (2012): total 8.9%, men 8.9%, women 9.0% (Gu 2018)
Europe (1992-2010, colon only): total 6%, men 4%, women 7% (Aleksandrova 2014)
Germany (2018): total 11.3% (Behrens 2018, figure 3)
Lebanon (2018): men 17%, women 10% (Charafeddine 2017)
New Zealand (2013-2014): total 4.4% (Richardson 2016)
UK (2015); total 4.6%, men 4.1%, women 5.1% (Brown 2018, Supplement F)
————————————————————————————————————————————
Physical inactivity is defined as lack of bodily movements produced by skeletal muscles that result in energy
expenditure. It is an established risk factor for colon cancer (Namasivayam 2017) and its definition includes
occupational, sports, conditioning, household or other activities (Caspersen 1985). It is inversely associated with
cardiorespiratory fitness (Kohler 2016, World Cancer Research Fund / American Institute for Cancer Research.
Continuous Update Project Expert Report 2018, accessed 28May20), but its relationship with rectal cancer is
inconsistent (Namasivayam 2017). Sedentary behavior (energy expenditure comparable to or less than sitting or
reclining), exemplified by prolonged TV viewing, is associated with an increased risk of colorectal cancer,
independent of exercise and obesity (Nguyen 2018, Schmid 2014).
Possible mechanisms for this association of physical inactivity with colorectal cancer, independent of excess weight,
include an increased inflammatory response from prolonged bed rest. This is demonstrated by increases in plasma C
reactive protein and IL6, decreases in IL10 and changes in other inflammatory markers (Bosutti 2008, Jurdana
2015). Physical inactivity is also associated with diet and smoking (Morris 2018) and lower Vitamin D levels
(Morales-Oyarvide 2016).

Page 4
Colorectal cancer due to excess weight
Excess weight (overweight), typically defined as a body mass index (BMI) of 25 or more (Centers for Disease
Control and Prevention, accessed 28May20), and obesity, defined as a BMI of 30 or more, are associated with a
30-70% increased risk of colon cancer in men, but the association is less consistent in women (Bardou 2013).
Abdominal obesity may be more important than overall obesity (Dong 2017). The terms overweight and obesity are
unfortunately used interchangeably in the literature.
The population attributable fraction of colonic and rectal cancer due to a BMI of 25 or more varies by region, gender,
site (colon versus rectum) and by study (see Table 4). These figures may be underestimates since average weight is
increasing worldwide (De Pergola 2013). We use 10-20% in Table 1 as the population attributable fraction because
the figures for the sole US study appear to be an outlier for Western countries and are substantially less than those
for Canada or North America in other studies.
————————————————————————————————————————————
Table 4 - Population attributable fraction of colorectal cancer due to excess weight
Region Men-colon Men-rectum Women-colon Women-rectum Source
Worldwide 13% 6% 8% 4% Arnold 2015
North America:
North America 21% 11% 11% 6% Arnold 2015
United States 5.1% (colorectum) 5.4% (colorectum) Islami 2018
Canada 20.5% 10.8% 9.7% 4.8% Zakaria 2017
Canada (Alberta) 12.2% (colorectum, men and women) Grundy 2017b
Europe:
Europe 15% 5% 7% 2% Aleksandrova 2014
Europe 16-19% 8-10% 9-11% 5% Arnold 2015
Germany 13.2% (colorectum, men and women) Behrens 2018
Germany 31.6% 11.3% 15.3% 2.7% Wienecke 2018
United Kingdom 15.3% (colorectum) 6.8% (colorectum) Brown 2018
Supp Table F
Australasia:
Australia 11.7% (colorectum) 5.5% (colorectum) Whiteman 2015
Australia 20.4% (colorectum) 4.0% (colorectum) Vajdic 2018
New Zealand 9.0% (colorectum, men and women) Richardson 2016
Other:
China 4.9% (colorectum) 5.8% (colorectum) Gu 2018
Korea 15.1% (colorectum) 22.5% (colorectum) Lee 2019
Lebanon 22% 19% 6% 3% Charafeddine 2017
Malaysia 4.5% (colorectum) 0.9% (colorectum) Naing 2017
Nigeria 7.6% (colon) 7.0% (colorectum) Odutola 2019
————————————————————————————————————————————
Excess weight directly promotes cancer via several chronic stressors, including chronic inflammation (typically low
grade, subclinical and affecting white adipose tissue), sex hormone biosynthesis and immune system dysfunction (De
Pergola 2013). First, regarding chronic inflammation, obesity promotes tumor growth through its association with
adipocyte hypoxia, which induces proinflammatory factors that create a microenvironment supportive of tumor growth
(Divella 2016, Bastard 2006, Quail 2019). Higher serum levels of C reactive protein are associated with increased
risk of colorectal cancer, but not other obesity related cancers (Izano 2016). Second, obesity causes elevated levels
of aromatase, the rate limiting enzyme in estrogen biosynthesis (Zahid 2016, Sato 2012), although estrogen can both
protect against and promote colon cancer (Chen 2012). Third, adipose tissue is considered an important and active
component of the innate immune response, which can create a favorable niche for tumor development in obese
individuals by secreting hormones, growth factors, adipokines (leptin and adiponectin) and proinflammatory cytokines
(Riondino 2014, Ahechu 2018). Of note, obesity is also associated with insulin resistance and abnormalities of the
IGF-I system (Avgerinos 2018).
Excess weight is also associated with physical inactivity (Sugiyama 2016), diet and type II diabetes, as discussed
elsewhere in this paper.

Page 5
Colorectal cancer due to tobacco
Tobacco smoking causes colorectal cancer and adenomas (The Health Consequences of Smoking—50 Years of
Progress: A Report of the Surgeon General (2014), page 204, PDF page 233). The population attributable fraction
of colorectal cancer due to smoking varies by country and gender (see Table 5). Biomarkers of smoking behavior
exhibit stronger associations with colorectal cancer than self-reporting, providing additional evidence for a role for
tobacco in this malignancy (Cross 2014).
————————————————————————————————————————————
Table 5 - Population attributable fraction of colorectal cancer due to tobacco
North America:
US (1997 deaths): men and women 12% (Chao 2000)
US (2011 deaths): adults age 35+: 9.7% of deaths, men 11.2%, women 8.0% (Siegel 2015-Table)
US (2014 cases): adults age 30+: total 11.7%, men 13.5%, women 9.7% (Islami 2018)
Canada (Alberta, 2012 cases): total 11.4% (Grundy 2017b)
Europe:
Europe (1992-2010 cases): total 4%, men 4%, women 1% (Aleksandrova 2014)
Germany (2008 cases): men 10%, women 5% (Wienecke 2014)
Norway (2002 cases): 12% of women (Gram 2009)
Australasia:
Australia (2010 cases): total 6.4%, men 5.9%, women 7.0% (Whiteman 2015)
China (2012 cases): total 4.9%, men 8.4%, women 0.4% (Gu 2018)
Korea (2012 deaths): men 4.3-7.8%, women 1.1-8.4% (Kong 2016)
New Zealand (2013-2014 cases): total 2.5% (Richardson 2016)
————————————————————————————————————————————
Cigarette smoking causes colorectal cancer via multiple mechanisms. First, cigarette smoke condensate and nicotine
cause epigenetic modification by increasing DNA methyltransferase 1 expression, which catalyzes the transfer of a
methyl group from S-adenosyl methionine to the 5’ position of cytosines in the cytosine-guanine dinucleotide
sequence (Xiao 2015). Cytosine methylation is a major epigenetic modification of DNA which influences a large
number of genes and many cellular processes (Phillips 2008). Cigarette smoke carcinogens may cause
hypermethylation of the APC 1A promoter (Barrow 2017), associated with serrated adenomas (Fu 2009, Figueiredo
2015), which may explain its association with MSI high, CIMP positive and BRAF mutation positive subtypes,
characteristic of the serrated neoplasia pathway of colorectal cancer (Drew 2017, LImsui 2010). The association of
smoking with synchronous colorectal tumors suggests it has a field effect (Drew 2017).
Second, cigarette smoking is associated with colorectal adenomas in general, not just serrated adenomas
(Shrubsole 2008). Cigarette smoke contains at least 60 carcinogens including ammonia, arsenic, benzopyrene,
cadmium and formaldehyde, as well as polynuclear aromatic hydrocarbons, heterocyclic amines, nitrosamines and
aromatic amines (IARC 2004). These carcinogens reach the colorectal mucosa by direct ingestion or through the
blood and may have a direct effect on the colon (Gram 2009). Adenomatous formation may also be mediated through
chronic inflammation (Ye 2004).
Third, smoking alters the gut microbiome (Lee 2018) including the risk of C. difficile infection (Rogers 2012). Tobacco
metabolites in the colonic mucus layer, including nicotine, may interact with intestinal bacteria and affect GI
secretions and motility (Kato 2010). In addition, tobacco related immunosuppression may alter bacterial flora, which
alters cancer risk, as discussed below.
Fourth, smoking may alter expression of cell cycle proteins, proapoptotic proteins, epithelial-mesenchymal transition
markers and cathepsin D expression, through an unknown mechanism (Kim 2017a).
Fifth, smoking may interact with genetic variants in spindle assembly checkpoint genes which can drive aneuploidy
and enhance colorectal cancer risk (Zhong 2015). The precise mechanism is unknown, although nicotine and
polycyclic aromatic hydrocarbons may induce genomic instability.
Finally, smoking status is associated with colorectal cancer risk factors of diet quality and obesity (Alkerwi 2017).
Colorectal cancer due to alcohol
Alcohol is associated with colorectal cancer and is a major carcinogen in general. Alcohol consumption is associated
with a modestly increased risk of cancer of the colorectum in men, to a greater extent than in women (Fedirko 2011,
Grundy 2016, Wang 2015). The association may be affected by race / ethnicity, lifestyle factors and alcoholic
beverage type (Park 2019).

Page 6
The attributable fraction of colorectal cancer cases due to alcohol varies from 4-13%, based on geography, site and
gender (see Table 6).
————————————————————————————————————————————
Table 6 - Population attributable fraction of colorectal cancer cases due to alcohol consumption
North America:
US (2014): total 12.8%, men 17.1%, women 8.1% (Islami 2018)
Alberta, Canada (2012): total 3.9%, men 5.4%, women 2.1% (Grundy 2016, Table 5).
Europe:
Europe (1992-2010): total 4% (colon 2%, rectal 6%), men 7% (colon 5%, rectal 12%), women 1% (colon NA, rectal
2%) (Aleksandrova 2014)
Australasia:
Australia (2010): colon 8.0%, rectum 11.8% (Pandeya 2015)
Australia (2010): total 9.0%, men 12.9%, women 4.2% (Whiteman 2015)
New Zealand (2013-2014): total 6.6% (Richardson 2016)
Other:
China (2012 cases): total 5.4%, men 8.7%, women 1.1% (Gu 2018)
Lebanon (2008): heavy drinkers: males 6%, females 0.2% (Charafeddine 2017)
Nigeria (2012-14): men 15.0%, women 2.3% (Odutola 2017)
————————————————————————————————————————————
Proposed mechanisms of action include: (1) ethanol consumption leads to microbial production of acetaldehyde,
which degrades folate (Homann 2000), which is protective against colon cancer; alcoholism is also associated with a
diet low in folate and fiber (Rossi 2018); (2) alcohol impairs the ability to absorb and break down vitamins A, B
complex, C, D, E and carotenoids (NIH > NCI > Alcohol and Cancer Risk, accessed 28May20); (3) alcoholic
beverages may contain carcinogens including nitrosamines, asbestos fibers (Fitzgerald 2019), phenols and
hydrocarbons; (4) chronic alcohol consumption produces genomic DNA hypomethylation, an early step in colonic
carcinogenesis (Choi 1999, Nishihara 2014); (5) alcohol activates liver procarcinogens by inducing cytochrome
P450 enzymes (Djordjević 1998); (6) alcohol consumption generates reactive oxygen species which can damage
DNA, proteins and lipids (He 2017), (7) alcohol increases blood levels of estrogen (Seitz 2007), (8) acetaldehyde and
other alcohol metabolites leads to immune modulatory effects (Rossi 2018).
Although the risk of head and neck cancers decreases after cessation of drinking (Marron 2010), the relationship at
other sites including the colon may be more complex (LoConte 2018).
Colorectal cancer due to diet
A proinflammatory diet (generally a Western diet high in red or processed meat, fats and sweets and low in
vegetables, fruits and dietary fiber), has been associated with an increased risk of colorectal cancer in US studies
involving health professionals (Tabung 2018a), women (Tabung 2015, Shivappa 2014a), retirees (Wirth 2015) and
a multiethnic cohort (Harmon 2017). Similar results were reported based on worldwide meta-analyses and reviews
(Shivappa 2017a, Jayedi 2018, Pan 2019) and in reports from Iran (Rafiee 2019, Shivappa 2018), Jordan
(Shivappa 2017b), Korea (Cho 2016a, Ryu 2019-women only) and Spain (Obón-Santacana 2019).
Other reports have focused on specific attributes of the Western diet. The World Cancer Research Fund and
American Institute for Cancer Research found that consuming red meat and processed meat increases the risk of
colorectal cancer (Diet, Nutrition, Physical Activity and Cancer: a Global Perspective 2018); these results were
recently summarized (Bernarba 2018). Associations may be stronger in men than women (Tabung 2017, Petimar
2018). Use of multivitamins or calcium reduces the risk of colorectal cancer (Erdrich 2015, Heine-Bröring 2015, Han
2015).
The attributable fraction of colorectal cancer cases due to consumption of red meat, processed meat and low dietary
fiber varies from 3% to 21% based on geography, dietary component measured and gender (see Table 7). For Table
1, we used an estimate of 5-10% for the population attributable fraction of colorectal cancer in the United States.
————————————————————————————————————————————
Table 7 - Population attributable fraction of colorectal cancer cases due to diet
North America:
US (2014):

Page 7
red meat: total 5.4%, men 6.6%, women 3.9%
processed meat: total 8.2%, men 10.3%, women 5.8%
low dietary fiber: total 10.3%, men 9.3%, women 11.3%
low dietary calcium: total 4.9%, men 3.9%, women 6.0% (Islami 2018)
Canada (Alberta, 2012):
excess red meat: total 9.5%, men 15.4%, women 1.8%
excess processed meat: total 2.9%, men 3.8%, women 1.6%
insufficient fiber: total 6.0%, men 6.6%, women 5.2%
inadequate vitamin D: total 9.2%, men 10.4%, women 7.8%
inadequate calcium: total 7.1%, men 5.8%, women 8.9% (Grundy 2017b)
Europe:
Europe (1992-2010): diet quality: total 5%, men 6%, women 2% (Aleksandrova 2014)
Denmark (2016-2045):
eliminating intake of red/processed meat: total 19.8%, men 24.4%, women 14.3%
reducing intake: total 9.1%, men 11.4%, women 6.4% (Lourenço 2018)
Germany (2018):
total red meat consumption: 0.7%
total processed meat consumption: 11.4%
total dietary fiber intake: 16.4%
total fruit and non-starchy vegetable consumption: 3.6% (Behrens 2018, figure 4)
UK (2010): red/processed meat: total 21.1%, men 24.8%, women 16.4% (Parkin 2011)
Australasia:
Australia (2010): red/processed meat: total 17.7%, men 21.8%, women 12.6% (Nagle 2015)
Australia (2010):
red/processed meat: total 17.6%, men 21.7%, women 12.5%
insufficient fiber: total 17.6%, men 18.6%, women 16.2% (Whiteman 2015)
China (2012):
red/processed meat: total 8.6%, men 9.1%, women 7.9%
low vegetables: total 17.9%, men 18.8%, women 16.8%
low fruit: total 6.4%, men 6.2%, women 6.7% (Gu 2018)
New Zealand (2013-2014):
red meat: total 4.8%, men 5.2%, women 4.4%
processed meat: total 2.6%, men 3.6%, women 1.6% (Richardson 2016)
Other:
Lebanon (2018): low adherence to Mediterranean diet: total 2%, men 2%, women 2% (Charafeddine 2017)
Columbia (2010):
red meat: total 12%, men 13%; women 10%
processed meat: total 14%, men 14%; women 13% (de Vries 2017)
————————————————————————————————————————————
Diet may promote colorectal cancer through multiple mechanisms that reinforce each other in unpredictable ways,
consistent with the behavior of complex systems. First, diet may promote systemic chronic inflammation, which is
central to colorectal carcinogenesis (Corley 2019), and may be mediated by inflammation associated cytokines that
drive cellular proliferation (Balkwill 2005). Higher concentrations of C reactive protein (Cho 2018, Zhou 2014, Izrano
2016) are associated with an increased risk of colon cancer. In support, the use of aspirin or other NSAIDs is
associated with a reduced risk of colon cancer, attributed to their anti-inflammatory properties (Wei 2009, Erdrich
2015).
Second, proinflammatory diets containing saturated fat and high glycemic index carbohydrate (potatoes, white bread
and short grain rice) but with low levels of fiber and omega 3 fatty acids may directly affect metabolic pathways,
leading to increased serum levels of insulin, triglycerides and nonesterified fatty acids, which may promote excessive
proliferation of colonic epithelial cells and potentially expose them to reactive oxygen species which damage DNA
(Bruce 2000). Similarly, foods with a high insulinemic potential (red and processed meat, margarine, poultry, French
fries, carbonated beverages) are associated with colon carcinoma (Tabung 2018b, Wang 2018).
Third, antioxidants and anti-inflammatory agents in food may mitigate the effects of occasional focal loss of the
colonic epithelial barrier, which promote focal proliferation and mutagenesis (Bruce 2000). Antioxidants in plant
extracts may directly inhibit lipid oxidation (Burri 2019). However, routine use of antioxidant supplements is not
recommended to prevent colorectal cancer, particularly in populations without significant nutrient deficiencies (Song
2015).

Page 8
Fourth, ingested carcinogens interact directly with colonic epithelial cells unless metabolized or absorbed in the small
intestine. The surface of well done red meat contains mutagenic substances including heme iron, heterocyclic amines
and polycyclic aromatic hydrocarbons (Gilsing 2013, Cascella 2018). Processed meats are also high in nitrates,
which may be converted to carcinogenic nitrosamines (Nagle 2015). Beef consumption and other high fat diets also
increase synthesis of fecal secondary bile acids, which have a carcinogenic effect on colonic epithelium via reactive
oxygen species and reactive nitrogen species, induction of DNA damage, mutation and apoptosis (Zeng 2015, Ajouz
2014).
Fifth, increased fiber consumption (non digestible carbohydrates) may increase fecal bulk, which dilutes carcinogens
from oxidative byproducts and decreases their bowel transit time and exposure to colonic epithelium (Grundy
2017a). Dietary fiber is fermented by colonic bacteria to yield butyrate, a short chain fatty acid and histone
deacetylase inhibitor that may protect against colorectal tumorigenesis by suppressing growth of tumor cells
(Bultman 2016, O'Keefe 2016) and influencing the microbiota profile (do Prado 2019). As noted in Table 7, low fiber
consumption by itself contributes from 6 to 18% of the attributable risk of colon cancer.
Sixth, diet has a fundamental role in colorectal cancer initiation, progression and prevention due to its influence on
gut microbiota composition (De Almeida 2019). The gut microbiome exists as a balanced, commensal community to
maintain homeostasis through symbiotic interactions with intestinal epithelium (Sheflin 2014). Alterations caused by
diet and infection (dysbiosis) can promote colorectal cancer (Gagnière 2016, Sun 2016). Specifically, subclinical
colorectal mucosal colonization with Escherichia coli and other Proteobacteria may be a risk factor for colorectal
cancer (Swidsinski 1998, Yang 2014, Jobin 2013). Alteration in bacteria flora may contribute to an expanded
community of “alpha bugs” (keystone pathogens) which harbor virulence traits that drive colon cancer development
(Hajishengallis 2012). The alpha bug hypothesis proposes that some microbiome members can remodel the colonic
bacterial community to enhance induction of alpha bugs, which co-opt other members of the microbial community and
may crowd out cancer protective microbial species (Sears 2011). Alpha bugs include E. coli harboring the polyketide
synthase (pks) island that encodes a putative genotoxin called colibactin (Arthur 2012, Eklöf 2017, Faïs 2018), and
enterotoxigenic Bacteroides fragilis (Wu 2009), Fusobacterium nucleatum (Rubinstein 2013, Liu 2018) and
Salmonella causing severe infections (Mughini-Gras 2018, Hernández-Luna 2019).
Persistent asymptomatic bacterial colonic infection may cause bacterial penetration of the inner mucus layer, inducing
increased permeability of tight junctions (Soler 1999, Grivennikov 2012), which delivers bacterial toxins directly to
the epithelium (Sears 2014). This may induce chronic inflammation and generate a pro-carcinogenic
microenvironment (Dejea 2013). In addition, inflamed epithelial cells under the stress of bacterial toxin exposure or
chronic bacterial infection generate reactive oxygen species and nitric oxide, which induce mutations (Dejea 2013).
These changes in microbiome, associated chronic inflammation and exposure to bacterial toxins induce local network
changes promoting dysplasia and ultimately malignancy (Lucas 2017, Montalban-Arques 2019). Dysplastic mucosa
is usually goblet cell depleted, lacks overlying mucus and has sparse underlying glycocalyx, which facilitates bacterial
contact with the mucosal surface to induce additional network and molecular alterations (Prorok-Hamon 2014).
Seventh, a proinflammatory diet may contribute to development of colorectal cancer by suppressing the adaptive anti-
tumor immune response (Liu 2017).
Eighth, human polymorphisms modulating the inflammatory response may affect microbiota influence on colorectal
cancer pathogenesis (Dejea 2013). In addition polymorphisms within DNA repair process genes can decrease their
efficiency and promote increased susceptibility to resident E. coli producing genotoxins (Buc 2013).
Association does not imply causation. Streptococcus gallolyticus subsp gallolyticus endocarditis is so strongly linked
to colorectal cancer (McCoy 1951, Klein 1997, Boleij 2013, Paritsky 2015) that its presence suggests the need for
colonoscopic examination. But S. gallolyticus gallolyticus is not a risk factor itself; instead, its virulence features
promote infections in colorectal cancer patients (Boleij 2011).
Colorectal cancer due to unhealthy lifestyle
Many studies have combined these behavioral risks to determine the reduction in population attributable fraction if all
are adapted. Overall attributable fractions range from 16% to 58%, as indicated in Table 8.
————————————————————————————————————————————
Table 8 - Population attributable fraction of colorectal cancer cases due to potentially modifiable risk factors
or unhealthy lifestyles
US (2014): men 58.2%, women 50.8%; risk factors are tobacco, alcohol, overweight, red / processed meat,
insufficient fiber, calcium, physical inactivity (Islami 2018)

Page 9
US (2015): women nurses 37%; risk factors are tobacco, alcohol, overweight, physical activity, multivitamins, calcium
intake (Erdrich 2015)
US (2012); white men 20%, white women 29%; risk factors are tobacco, alcohol, overweight, physical activity (Song
2016)
Europe (1992-2010): total 16%, men 22%, women 11%; risk factors are tobacco, alcohol, overweight, physical
inactivity, diet quality (Aleksandrova 2014)
————————————————————————————————————————————
Colorectal cancer due to germ line variations
The attributable fraction for non-advanced and advanced colorectal neoplasia is only 2-4% for family history
(colorectal cancer in a first degree relative, Hoffmeister 2010), although 16% to 35% of young US colorectal cancer
patients have an identifiable hereditary cancer syndrome (Pearlman 2017, Mork 2015). Although well described
cancer syndromes including Lynch syndrome and familial adenomatous polyposis confer a high risk of colorectal
cancer, they account for less than 5% of all cases (Jasperson 2010). However, it is estimated that 30% to 80% of
colon cancers may be due to less penetrant susceptibility genes (Jasperson 2010, Fuchs 1994, Strum 2019).
Lynch syndrome is an autosomal dominant familial colorectal cancer syndrome due to germ line mutations in
mismatch repair (MMR) genes MLH1, MSH2, MSH6 and PMS2 or in EPCAM. It is the most common known cause of
hereditary colorectal cancer and accounts for 4% to 13.5% of patients with early onset cancer (Pearlman 2017). The
MMR system maintains genomic stability by correcting errors (single base mismatches and insertion deletion loops)
of DNA replication. Mutations in hMSH2 and hMLH1 cause up to 90% of cases; mutations in hMSH6 cause 10% of
cases and mutations in hPMS2 are rare. Germ line deletions in EPCAM are found in a subset of families with a Lynch
syndrome phenotype but no MMR gene mutations, and may account for 6% of Lynch syndrome cases (Jasperson
2010).
Familial adenomatous polyposis (FAP), also autosomal dominant, is due to germ line mutations in APC, which
encodes a tumor suppressor part of the WNT signaling pathway. It is the second most common inherited colorectal
cancer syndrome and features hundreds to thousands of colonic adenomas, beginning in early adolescence. Almost
all untreated individuals develop colorectal cancer, typically by age 39 years, but occasionally (7%) by age 21.
Attenuated FAP is a less severe form characterized by an average of only 30 colonic adenomas and a lifetime risk of
colorectal cancer of 69% (Jasperson 2010).
MUTYH polyposis is caused by biallelic mutations in MUTYH (MYH), whose gene product is part of the base excision
repair pathway, which prevents G:C to T:A transversions caused by oxidative stress to highly mutagenic DNA bases.
Adenomatous polyps predominate but unlike attenuated FAP, hyperplastic polyps are common (Nielsen 2012).
Peutz-Jeghers syndrome (PJS) and juvenile polyposis syndrome (JPS) are hamartomatous polyposis conditions
associated with an increased risk of colorectal and other malignancies. Peutz-Jeghers syndrome is caused by germ
line mutations in STK11 (LKB1) and patients have a 81% to 93% lifetime risk of cancer, including an almost 70% risk
of GI cancer. Juvenile polyposis syndrome is caused by mutations in SMAD4 or BMPR1A, and features multiple
juvenile polyps, most prominently in the colon but also in the stomach and small bowel. Juvenile polyposis syndrome
patients have a lifetime risk of 39% for colorectal cancer (Jasperson 2010). In contrast, Cowden syndrome is a
hamartomatous polyposis that is not associated with an increased risk of colorectal cancer.
Cells have robust processes to limit errors during cell division, with at least 169 enzymes participating in DNA repair
or influencing DNA repair processes (Wood 2014). Half of these enzymes are directly involved in base excision
repair, nucleotide excision repair, recombinational repair, mismatch repair and direct reversal. These processes
reduce the transcription error rate by up to 99.9%, from one per 100,000 base pairs (Pray 2008) to one per 10-100
million base pairs (Yamada 2010). This repair process limits changes to protein structure and function that might
disturb existing network interactions or create new ones. Coupled with existing chronic stressors (risk factors), germ
line variations in these repair genes affect network stability and promote transitions to more unstable network states
associated with neoplasia.
Colorectal cancer due to random chronic stress or bad luck
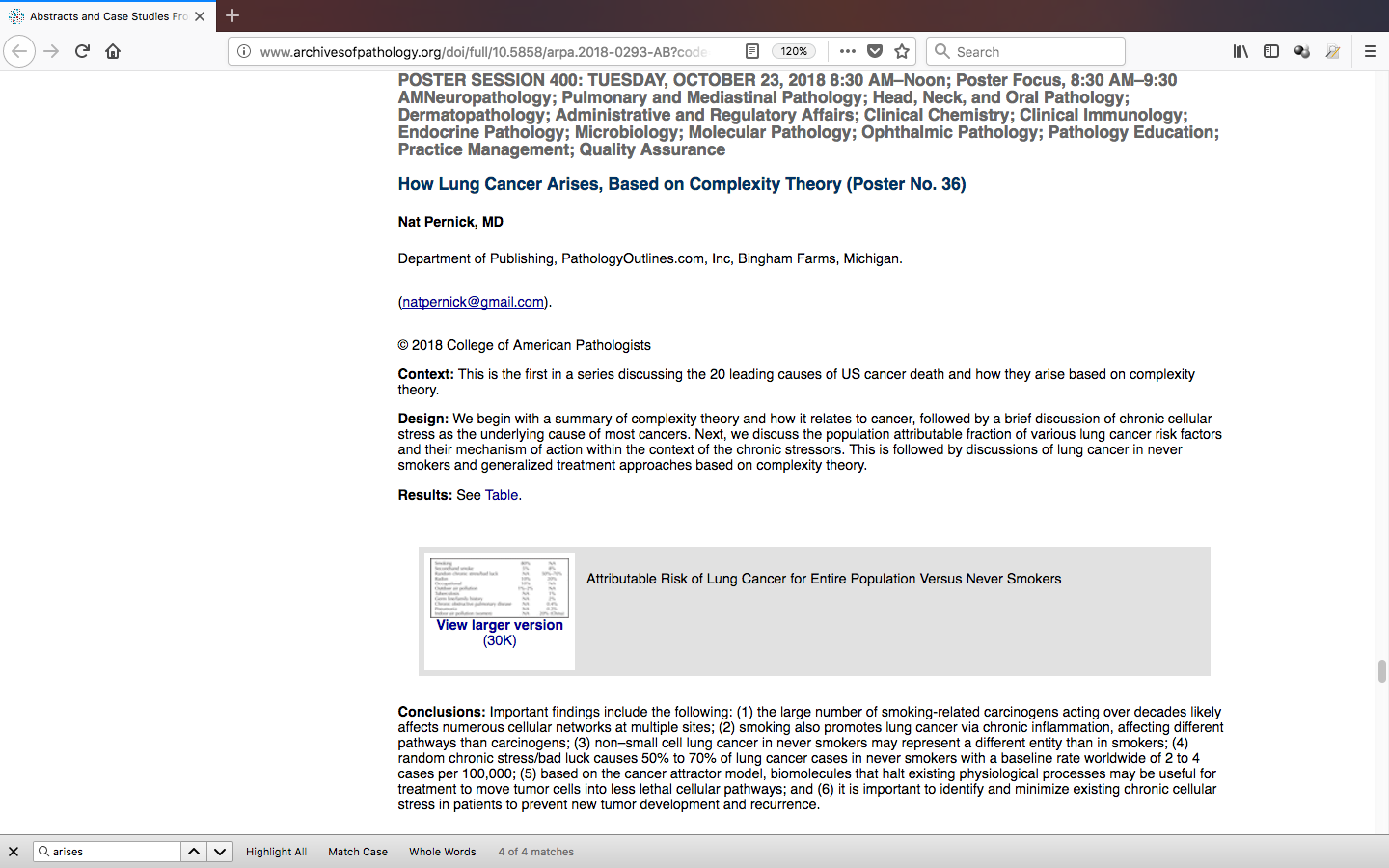
We previously attributed a surprising 50-70% of lung cancer cases in never smokers in North America and Europe to
random chronic stress or bad luck (Pernick 2018b-Session 400, poster 36, Abstract, Poster), although
ascertainment is made difficult by the overriding presence of tobacco use in causing lung cancer. For colon cancer,
the attribute risk of random chronic stress or bad luck appears to be <5% of cases, based on a calculation of 100%
minus the attributable risk of the known risk factors described above; of primary importance are the behavioral and
other factors listed in Table 1 (Song 2016). However, this calculation is not precise because PAFs may not be
additive due to the possible lack of independence of risk factors (Islami 2018)

Page 10
Risk factors with no estimates of population attributable fraction
Risk factors for colorectal cancer with population attributable fractions less than 1% or which have not been
calculated are described below.
Colorectal cancer due to aging
Advancing age is an important risk factor for colorectal cancer; 78% of cases are diagnosed at age 55 years and
older, and 15% occur at ages 45 to 54 years (Colorectal Cancer Screening (PDQ®) Health Professional Version,
accessed 29May20). The median age at diagnosis is 68 years (NIH > NCI > Age, accessed 29May20). However, the
population attributable fraction for aging has not been determined.
Aging acts in several ways to promote carcinogenesis. First, aging is associated with DNA methylation in normal
colonic tissue, particularly in patients at high risk (Wang 2020), that progresses to hypermethylation in colorectal
cancer (Ahuja 1998). These changes lower the threshold for malignant transformation (Nakagawa 2001, Xu 2014),
perhaps by altering chromatin conformation and stability (Lin 2015). They may also affect the mRNA expression of
adenoma and colorectal cancer related control genes (Galamb 2016).
Second, aging provides more time for chronic stressors to exert their effects (Martincorena 2015), which may take
decades. In addition, longer lifetimes allow for more interactions between the networks which they alter.
Third, aging is associated with immune system dysfunction, a known chronic stressor which causes malignancy
(Zhang 2016, Sadighi 2018). It may also affect the microenvironment (Marongiu 2016).
The rate of aging is controlled in part by nutrient sensing pathways (insulin or IGF1 signaling, mTOR, AMPK and
sirtuins) that have been evolutionary conserved from worms to humans (Yokoyama 2015). These pathways are also
commonly involved in carcinogenesis and cancer metabolism. Metformin, resveratrol, Rhodiola and other agents that
target these pathways may have both anti-aging and preventative, anti-colon cancer efficacy (Nguyen 2009,
McCubrey 2017)
Colorectal cancer due to type II diabetes
Type II diabetes is associated with a mildly elevated relative risk (1.2 to 1.4) of colorectal cancer (Tsilidis 2015, Lin
2014), particularly in men (Ma 2018), although a population attributable fraction risk has not been calculated. It is
unclear whether this association is due to shared risk factors (excess weight, diet, physical inactivity, aging) or due to
metabolic derangements of diabetes (hyperglycemia, insulin resistance, hyperinsulinemia, Giovannucci 2010) and
alterations of the insulin-like growth factor system (Scappaticcio 2017, Pollak 2004), which may create a low grade
chronic inflammatory state (Gristina 2015) and otherwise drive cancer metabolic reprogramming (Tudzarova 2015).
Colorectal cancer due to inflammatory bowel disease
Patients with longstanding ulcerative colitis and Crohn disease may have an increased risk of colorectal cancer which
increases with the duration and extent of colitis, degree of inflammation, the presence of primary sclerosing
cholangitis and family history of colorectal cancer (Triantafillidis 2009, Laukoetter 2011). No population attributable
fraction has been calculated. However, the incidence has been decreasing in western countries (Kim 2014) and there
is substantial heterogeneity between studies; some authors question whether there actually is an increased risk
(Wheat 2016, Adami 2016).
Although the pathogenesis of colorectal cancer in these patients is poorly understood, there are several suggested
mechanisms. First, genetic changes and genomic instability in inflammatory bowel disease and the adenoma-
carcinoma sequence may be similar; in both, histology progresses from no dysplasia to indefinite dysplasia, low
grade dysplasia, high grade dysplasia and finally to invasive adenocarcinoma, although steps can be skipped
(Triantafillidis 2009). Dysplastic lesions tend to be multifocal, in contrast to non-IBD, non-germ line cases, indicating
a field effect (Ullman 2011). These changes may be due to establishment of new hierarchies of molecular patterns,
which may lack associated histologic changes (Pernick 2018c).
Second, host inflammation affects several other risk factors, including the composition and functional capabilities of
gut microbiota (Arthur 2013). In addition, inflammatory mediators TNF, IL17A and IL23 and byproducts such as
reactive oxygen and nitrogen species produce genetic and epigenetic modifications that may lead to carcinogenesis
(Arthur 2013, Däbritz 2014). Hypermethylation of the CpG island silences tumor suppressor genes, including DNA
repair genes such as hMLH1 (Kim 2014).
Colorectal cancer risk reduction due to low dose aspirin
Low dose aspirin is recommended by the US Preventive Services Task Force for the primary prevention of
cardiovascular disease (CVD) and colorectal cancer in adults aged 50 to 59 years who have a 10% or greater 10
year CVD risk, are not at increased risk for bleeding, have a life expectancy of at least 10 years and are willing to
take low dose aspirin daily for at least 10 years (US Preventative Services Task Force, accessed 30May20, but see
Chubak 2015). In Australians, daily aspirin use may reduce the incidence of colorectal cancer by 2.3% in men and

Page 11
2.0% in women (Whiteman 2015, Wilson 2015). The Cancer Council Australia recommends aspirin use for Lynch
Syndrome patients, and suggests its use for all people aged 50-70 years at average risk of colorectal cancer who
have a life expectancy of at least 10 years (Cancer Council Australia, Clinical Guidelines Network, accessed
30May20).
Colorectal cancer risk reduction due to menopausal hormone therapy
Menopausal hormone therapy but not endogenous estrogen exposure (Neumeyer 2018), appears to reduce the risk
of colorectal cancer (Mørch 2016, Botteri 2017, Lin 2012), possibly due to receptors for follicle stimulating hormone
and luteinizing hormone expressed in colorectal tumors (Marlicz 2018). However, it may cause more cancer in the
breast than it prevents in the colon (Whiteman 2015).
Colorectal cancer due to asbestos exposure
Whether asbestos exposure is associated with colorectal cancer incidence or mortality is controversial (IARC
Monogr Eval Carcinog Risks Hum 2012, Paris 2017).
Treatment strategies for colorectal cancer based on complexity theory
Treatment of colorectal cancer is based on stage (Colon Cancer Treatment (PDQ®)–Health Professional Version,
accessed 30May20) and consists of surgery and chemotherapy for nonmetastatic disease. For metastatic disease, it
includes targeted therapy, which acts on mutations in the EGFR and BRAF pathways (Ducreux 2019). For all stages,
the 5 year relative survival rate is 63% for colon cancer and 67% for rectal cancer (Cancer Facts & Figures 2020). It
increases to 90% for localized colorectal cancer, although only 39% of patients have localized disease.
Current colorectal cancer treatment is based on reductionist principles, namely killing tumor cells where they exist,
and assuming that is curative. Reductionist thinking is helpful to understand how premalignant lesions arise from a
small number of local factors. However, advanced or disseminated disease is likely due to an altered systems biology
(Koutsogiannouli 2013), with changes in networks throughout the body that typically will not revert to normal by
destroying or removing the original tumor.
We propose that local changes, over long periods of time, create instabilities in adjacent networks that ultimately may
propagate throughout the body. To attain high cure rates, we recommend treatment that targets as many networks as
possible. It is likely that many of these factors are not additive, but multiplicative, so a multi-pronged approach is
particularly important. It should include targeting: (1) tumor cells - kill by multiple, distinct methods to compensate for
the diversity of risk factors and their widespread effects; (2) tumor cells - move from "cancer attractor" networks
towards more differentiated or less hazardous states; (3) tumor microenvironment - target multiple aspects which
nurture the tumor, (4) tumor associated immune system dysfunction - identify and counter, (5) chronic stressors active
in this patient - identify, reduce and mitigate, (6) premalignant lesions - eliminate through more effective screening, (7)
germ line changes - identify and target those associated with tumor promotion and (8) overall patient health - promote
to detect disease earlier and expand treatment options
Of course, treatment strategies are also affected by the degree to which existing disease is an immediate threat to life
versus a risk for later recurrence, metastases or new tumors. In addition, trial and error may be necessary to
determine which treatment strategies are synergistic or antagonistic.
1. Curative therapy requires killing tumor cells by attacking as many targets as possible in their disordered
networks via distinct mechanisms. A diverse treatment approach is important for several reasons. First, colorectal
cancer is inherently more difficult to treat than many childhood tumors. Developing curative treatment for childhood
leukemia, Hodgkin lymphoma and testicular cancer involved refining combinations of agents with proven impact
against tumors, typically through varied mechanisms of action and by minimizing adverse side effects (Mukherjee:
The Emperor of All Maladies, 2010, National Cancer Institute-The "Accidental" Cure, accessed 30May20). Yet
these cancers typically affect the young, have no prominent risk factors and show no field effect. In contrast,
colorectal cancer has a high median age (colon cancer: 68 years in men, 72 years in women; rectal cancer: 63 years
in men and women, Colorectal Cancer Facts & Figures 2017-2019) and patients often have multiple risk factors
which generate field effects: excess weight, physical inactivity, Western diet, tobacco and alcohol use and germ line /
family history. Thus, although risk factors for colorectal cancer may produce less tumor heterogeneity than tobacco
does for lung cancer (Pernick 2018b, Pernick 2018c, Govindan 2012), curative therapy must combat a
heterogeneous tumor with multifocal growth.
Second, treatment should attack multiple molecular targets within tumor networks. Currently, treatment is usually
based on the predominant molecular driver mutation (Inamura 2018). Yet colorectal cancer often exhibits molecular
heterogeneity within the same patient (Punt 2017, López 2017) and molecular features may evolve over time
(Baretti 2018).
Third, curative therapy must target the effects of tumor cells on other cells. Tumor cells not only directly damage
tissue and replace other tumor cells killed by treatment, but they may produce cytokines that assist tumor cell survival

Page 12
and proliferation (exemplified by Hodgkin Reed-Sternberg cells, Mani 2009), and may have a prion-like effect of
inducing malignant behavior in neighboring cells (Ann Bom 2012). Acknowledging that treatment typically will not kill
100% of tumor cells, it is important to counter properties of tumor cells that survive.
In addition to current chemotherapy which attacks growth related networks and specific mutations, we suggest
treatment which takes advantage of the sophistication present in physiologic cells but lost in the somewhat chaotic
cancer cells of advanced and aggressive colorectal cancer. This includes “lethal challenges” that require
sophisticated functioning for cells to survive, such as high dose methotrexate with leucovorin rescue (Howard 2016),
which might be adapted to colonic cancer cells.
2. Curative therapy should move malignant networks towards less lethal states. Kauffman discovered that a
complex network of thousands of mutually regulating genes in normal cells may produce a stable equilibrium state
called an attractor which corresponds to gene expression profiles specific to each cell type (Kauffman 1969; see also
Noble 2015). Attractors have been analogized to a low energy state or valley on a topographic diagram that pulls in
cells with similar network configurations (Waddington, The Strategy of the Genes, 1957, Figure 1). Attractors also
maintain cellular network stability against common perturbations. Cancer attractors may arise when chronic cellular
stress overrides this stability to alter cellular networks, producing new intermediate states and malignant cells that
exhibit relatively stable gene expression profiles but with malignant properties. These phenotypic changes may not be
due to mutations; after all, 200+ cell types exist based on the same genome but with differences in expression of
gene regulatory networks (Huang 2009). Cancer attractors may also exist in healthy genomes but are normally not
accessible, analogous to dangerous cliffs that exist but are avoided by well planned highways (Huang 2009).
Changes to the gene expression profile during carcinogenesis provides a means to access these dangerous and
unstable cellular states.
Cancer attractors may be “close” to developmentally immature states (Huang 2009). As a result, tumor cells may
harness the molecular cues used to pattern an embryo to enhance growth, recruit stromal cells and coordinate
spread (Aiello 2016), explaining why oncogenesis recapitulates ontogenesis (Huang 2009). Network altering
treatments cannot typically move tumor cells or premalignant cells back to their physiologic state, but they can push
them towards alternative, more differentiated phenotypes with reduced malignant properties. This may explain the
powerful anti-tumor effects of differentiation agents such as retinoids for acute promyelocytic leukemia and childhood
neuroblastoma (Nowak 2009), progestin for endometroid intraepithelial neoplasia (Wong 2019) and other lineage
reprogramming agents (McClellan 2015, Gong 2019).
A theoretical framework to move malignant networks to a less hazardous state has been described (Huang 2013,
Kim 2017b). However, the dynamic nongenetic heterogeneity of tumor cells makes them moving targets and drives
replenishment of the tumor with surviving, non-responsive cells. Although the nonlinear functioning of gene regulatory
networks makes prediction difficult, it has been suggested that network rewiring could be accomplished with constant
perturbation of networks (Cho 2016b).
3. Curative therapy should target different aspects of the microenvironment nurturing the tumor. Chronic
stressors produce a microenvironment which may nurture mutated cells, steer cellular networks towards malignant
pathways (Mbeunkui 2009, Morgillo 2018), help them escape immune surveillance (Dalgleish 2006) and ultimately
promote invasion by activating cells to mimic physiologic “invasion” of wounded epithelium through the extracellular
matrix (Bleaken 2016, Coussens 2002). As a result, it may be important to interfere with the complex crosstalk
between cancer cells, host cells and the extracellular matrix (Sounni 2013, Carvalho 2019, Crawford 2017) by
normalizing aberrant properties (Gkretsi 2015) and by disrupting the fertile “soil” necessary for the cancer “seeds” to
grow (Fidler 2003). Normalizing the microenvironment may enhance drug delivery and effectiveness (Polydorou
2017, Stylianopoulos 2018) or make existing tumors or intermediate states more susceptible to immune system
attack or other treatments.
To rewire these malignant networks, we suggest treatments based on emulating pathways which halt processes
associated with malignancy, such as rapid cell division in embryogenesis (Kermi 2017, Jukam 2017), chronic
inflammation, wound healing (Sulniute 2016, Shah 2018, Kareva 2016) or liver regeneration (Mao 2014, Abu
Rmilah 2019) We speculate that these halting factors are likely to impact malignant pathways in colorectal and other
cancers. In addition, applying environmental stressors that promote the activation of multicellular programs may push
cells towards the compensatory inactivation of unicellular processes relied on by cancer cells, which may render them
unviable (Trigos 2018).
4. Curative therapy should counter immune system dysfunction that would otherwise limit tumor growth.
Cells evade the immune system through “camouflage and sabotage” as they acquire malignant characteristics
(Poschke 2011), resulting in tumor escape from immune surveillance (Ostrand-Rosenberg 2008). The dynamics of
this coevolutionary process may be similar to untreated HIV infection and CD4+ T cells, in which “escape mutants” of
HIV arise faster than the immune system can respond (Nowak 1995a, Goulder 1997).

Page 13
Immunotherapy has been successful for colorectal cancer through the use of checkpoint inhibitors in patients with
mismatch repair colorectal cancer, which has a relatively high mutational burden (Lynch 2016, Emambux 2018).
However, this reductionist approach focuses on only one aspect of one immune system pathway - a broader strategy
may be needed for curative therapy.
5. Curative therapy requires eliminating or reducing the chronic stressors which cause network changes that
ultimately lead to cancer. These chronic stressors act through varied mechanisms and reinforce each other in
unpredictable ways. Their interactions create chronic inflammation and carcinogens that exert independent effects
and promote progression towards malignancy. As the magnitude of these chronic stressors is reduced, the
interactions are also reduced and networks may revert towards more stable states.
Reducing chronic stressors may not affect the existing tumor but should reduce the incidence of new tumors. Most
chronic stressors for colorectal cancer are behavioral. However, we suggest that it may be helpful to detect and
counter the inflammatory process in general, because (a) some patients may not change their behavior, (b) some
chronic stressors are not behavioral and (c) advanced network changes may not be reversible even when behavior
does change.
6. Curative therapy for colorectal carcinoma requires optimal detection and elimination of premalignant
lesions through more effective screening. Lack of screening, which finds and removes adenomas, is the major
risk factor for colorectal cancer, as described above. Any systemic plan to cure colorectal cancer must optimize all
steps in the prevention, detection and treatment process, which includes improving screening and overcoming
missed colorectal lesions after colonoscopy (Lee 2017).
7. Curative therapy requires targeting germ line changes associated with tumor production. Syndromes
associated with an elevated risk of colorectal cancer include Lynch syndrome, which impairs DNA repair (NIH > NCI >
Genetics of Colorectal Cancer, accessed 31May20) and PTEN hamartoma tumor syndrome, which damages
immune response (Eissing 2019). Although fixing these genetic errors does not seem feasible in the near future,
chemoprevention strategies for high risk patients are under investigation, including non-steroidal anti-inflammatory
drugs, polyamine inhibitors, the antihelminth drug mebendazole (Williamson 2016, Gerner 2018) and mTor inhibitors
(Yehia 2019).
8. Curative therapy becomes more likely when supported by efforts to optimize patient health. Improved
patient health may not only reduce chronic stressors which promote malignancy, but symptoms may be more obvious
in those with fewer health problems. In addition, better health increases the possibilities of effective treatments not
available to those with poor performance status or severe comorbidities.
Summary
Most important risk factors for colorectal cancer are preventable by behavioral changes, suggesting that dramatic
changes in death rates are possible with current treatment and preventive efforts. For existing tumors, the diverse
pathways of colorectal carcinogenesis suggest that no single treatment modality is likely to be curative. As aggressive
tumors and widespread disease are accompanied by systemic changes different in character from those in tumor
cells, we suggest combining multiple treatment strategies to attain cure: (1) kill tumor cells via multiple, distinct
methods to compensate for the diversity of risk factors and their widespread tissue effects; (2) move tumor cells from
"cancer attractor" network states towards more differentiated or less hazardous states; (3) target different aspects of
the microenvironment nurturing the tumor, (4) counter tumor associated immune system dysfunction, (5) identify,
reduce and mitigate patient related chronic stressors, (6) eliminate premalignant lesions through more effective
screening, (7) identify and target germ line changes associated with tumor promotion and (8) promote overall patient
health.

{kind=link}
{kind=link}
{kind=link}